LYMPHÔ B
Sở dĩ được gọi là lymphô bào B vì ở loài chim các lymphô bào này được tăng sinh và biệt hóa tại Bursa Fabricius (túi Fabricius). Người ta không tìm thấy một cơ quan nào tương đương với túi Fabricius ở loài có vú. Ngày nay người ta đã biết rằng, ở loài có vú các lymphô B được tăng sinh và biệt hóa ngay tại tủy xương. Lymphô B là tiền thân của các tương bào sản xuất ra kháng thể.
Sự biệt hóa của lymphô B
Ta có thể chia sự biệt hóa của lymphô B làm hai pha: pha biệt hóa độc lập với kích thích KHÁNG NGUYÊN lạ và pha biệt hóa phụ thuộc kích thích KHÁNG NGUYÊN lạ
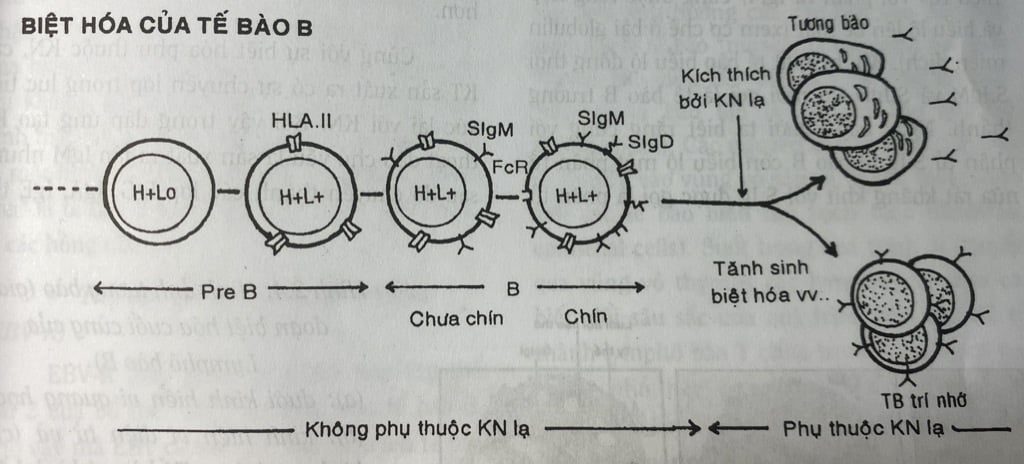
Biệt hoá độc lập với kháng nguyên lạ
Biểu hiện đầu tiên của sự biệt hóa thành dòng tế bào B là sự sắp xếp lại các gian nhỏ giữa các nhóm gien V, D, J để hình thành gien của chuỗi nặng rồi chuỗi nặng H (H) được tổng hợp và có mặt trong bào tương. Ở giai đoạn này tế bào được gọi dưới tên tế bào pré B. Sau đó là đến lượt sắp xếp lại các gian nhỏ V, J của chuỗi nhẹ L và tổng hợp chuỗi nhẹ. Khi có cả chuỗi nặng u và chuỗi nhẹ, phân tử IgM được hình thành và biểu lộ lên bề mặt tế bào tức là các IgM bề mặt (Surface IgM = S.IgM hay còn viết m.IgM; m=membrane). Các S.IgM này đóng vai trò thụ thể KHÁNG NGUYÊN của tế bào B. Lúc bề mặt chỉ mới biểu lộ riêng S.IgM thì tế bào B vẫn còn chưa trưởng thành (immature B cells). Nếu tiếp xúc với KHÁNG NGUYÊN ở giai đoạn chưa trưởng thành thì tế bào có thể bị bất hoạt hay loại trừ thay vì tăng sinh và biệt hóa nếu tiếp xúc khi đã trưởng thành. Tiếp theo IgM phân tử ICD (có cùng đặc hiệu KHÁNG NGUYÊN với phân tử IgM) cũng được tổng hợp và biểu lộ lên bề mặt (xem cơ chế ở bài globulin miễn dịch). Khi bề mặt tế bào biểu lộ đồng thời SIgM và S.Ign thì mới gọi là tế bào B trưởng thành. Ngày nay người ta biết rằng cùng với phân tử S.Ig tế bào B còn biểu lộ một phân tử nữa rất khăng khít với S.Ig được gọi là phân tử Igαβ (cấu trúc gồm 2 chuỗi α và β) giữ cho năng truyền tín hiệu từ S.Ig được kích thích – KHÁNG NGUYÊN đặc hiệu vào trong tế bào (tương đương như phân tử CD3 của TCR). Cụm 2 phân tử S.Iợ và IgxB được gọi là thụ thể của tế bào B (BCR = R Cell Receptor). Từ đây sự biệt hóa tiếp tục phải cần có KHÁNG NGUYÊN lạ kích thích.
Biệt hoá phụ thuộc với kháng nguyên lạ
Những tế bào B trưởng thành này nếu gặp KHÁNG NGUYÊN tương ứng đồng thời nhận được các tín hiệu giúp đỡ từ tế bào T thì sẽ tiếp tục biệt hóa. Ở bước biệt hóa này, các tế bào B sẽ tăng sinh và chia thành hai nhóm: hoặc trở thành các tương bào (là các tế bào tổng hợp và sản xuất ra KHÁNG THỂ có tính đặc hiệu tương ứng với KHÁNG NGUYÊN đã kích thích) ) hoặc trở thành những tế bào B có trí nhớ (memory B cells) cũng đặc hiệu với KHÁNG NGUYÊN. Các tế bào B có trí nhớ này sẽ giúp cho cơ thể khi tiếp xúc lại với KHÁNG NGUYÊN tương ứng có đáp ứng nhanh hơn, mạnh hơn và hiệu quả cao hơn.
Cùng với sự biệt hóa phụ thuộc KHÁNG NGUYÊN, các KHÁNG THỂ sản xuất ra có sự chuyển lớp trong lúc tiếp xúc lại với KHÁNG NGUYÊN. Bởi vậy trong đáp ứng tạo KHÁNG THỂ thoạt đầu chủ yếu là sản xuất ra lớp IgM nhưng sau đó chuyển thành các lớp IgG, IgA, IgE tùy thuộc vào điều kiện cụ thể. Điều đặc biệt cần ghi nhớ là tất cả các KHÁNG THỂ dù được chuyển sang lớp nào đều giữ tính đặc hiệu của IgM ban đầu. Phần này sẽ được giải thích trong bài nói về Globulin miễn dịch.
Các dấu ấn của lymphô B
Globulin miễn dịch bề mặt (S.Ig: Surface immunoglobuline)
Các tế bào B trưởng thành một phần hay hoàn toàn đều có globulin miễn dịch trên bề mặt. Các phân tử globulin miễn dịch bề m. (S.Igs) đóng vai trò làm thụ thể KHÁNG NGUYÊN của tế bào B. Các tế bào pré B, các tương bào chưa có hoặc không còn globulin miễn dịch ở bề mặt. Bởi vậy ta có thể nhận diện tế bào B nhờ dấu ấn S.Igs.
Thụ thể Fc (FcR: Fc Receptor)
Thụ thể Fc (FcR) hay phân tử CD16 là thụ thể của phần Fc của phân tử IgG. Ngoài tế bào B ta còn thấy FcR trên một số tế bào khác như các tế bào đơn nhân, tế bào NK. Ta có thể nhận biết tế bào có thụ thể Fc bằng kỹ thuật tạo hoa hồng với hồng cầu cừu đã mẫn cảm trước với kháng thể IgG chống hồng cầu cừu. Dưới kính hiển vi quang học các tế bào có FcR sẽ tạo ra một hình ảnh giống như hoa hồng mà “nhụy hoa” là tế bào lymphô B có FcR với “cánh hoa” là các hồng cầu cừu.
Thụ thể với Epstein- Barr virus
EBV-R (hay phân tử CD21 hay thụ thể thứ 2 của bổ thể: CR2) có mặt trên tế bào B. Nhờ vậy mà EBV có thể gắn đặc hiệu lên tế bào B. Nhiễm EBV vào tế bào B có thể làm cho tế bào này trở thành bất tử (immortalization) và liên tục nhân lên và trong những điều kiện nhất định có thể trở thành ung thư.
Phân tử nhóm phù hợp mô lớp II
Tế bào B và các tế bào có chức năng trình diện KHÁNG NGUYÊN như các loại đơn nhân thực bào có biểu lộ thường trực trên bề mặt các phân tử của nhóm phù hợp mô lớp II (ở người là phân tử HLA- DR). Các phân tử HLA- DR này ở tế bào B cũng có chức năng trình diện KHÁNG NGUYÊN cho tế bào T nhất là trong đáp ứng miễn dịch thì hai.
LYMPHÔ T
Sự trưởng thành của lymphô T
Một nhóm tế bào gốc dòng lymphô được di chuyển đến Thymus và tại đây chúng được tăng sinh, biệt hóa và giáo dục để trở thành lympho T trưởng thành. Nguồn gốc của Thymus được hình thành từ túi hầu (pharyngeal pouches) thứ 3 và 4 phát triển xuống trung thất trước tim. . Thoạt đầu, các tiến Thymô bào (Thymocytes) đến vùng vỏ ngoài của tuyến, tại đây dưới ảnh hưởng của các liên bào đặc biệt được gọi là tế bào “nurse” (nurse cells) các tiền Thymô bào chịu các thay đổi và tăng sinh để cho các tế bào lớn là nguyên bào lymphô (lymphoblast). Các nguyên bào lymphô dần dần di chuyển vào vùng vỏ sâu và đi qua một mạng lưới các tế bào biểu mô bạch tuộc (dendritic epithelial cells). Suốt trong quá trình di chuyển qua vùng vỏ thymus các lymphô bào chịu các biến đổi sâu sắc của quá trình biệt hóa để trở thành lymphô bào T chưa trưởng thành có kích thước nhỏ hay trung bình đi vào vùng tủy thymus. Ở tủy Thymus, các lymphô bào tiếp xúc với một mạng liên bào thưa hơn và xen lẫn trong đó có các tế bào xòe ngón tay (interdigitating cells) thuộc nhóm đơn nhân thực bào để trở thành tế bào T trưởng thành đi vào tuần hoàn. Tại Thymus các lymphô bào được nhân lên rất nhanh chóng song cũng rất nhiều (có thể đến >95%) chết tại chỗ – đây là hậu quả của sự chọn lọc hay còn gọi là quá trình giáo dục ở Thymus đối với tế bào T.
Sự giáo dục, chọn lọc tế bào T tại Thymus
Quá trình biệt hóa và trưởng thành từ những tế bào gốc dòng lymphô để trở thành các lymphô T có chức năng xảy ra tại Thymus. Những tế bào T trưởng thành phải có khả năng nhận diện và phản ứng lại các KHÁNG NGUYÊN lại được trình diện trong nhóm phù hợp mô nhưng lại không được phản ứng với KHÁNG NGUYÊN của bản thân. Để thu được các thuộc tính trên các lymphô bào T phải “học hỏi và chịu sự chọn lọc khắc nghiệt , bị hủy diệt hay bất hoại nếu không đạt các “tiêu chuẩn” nêu trên.
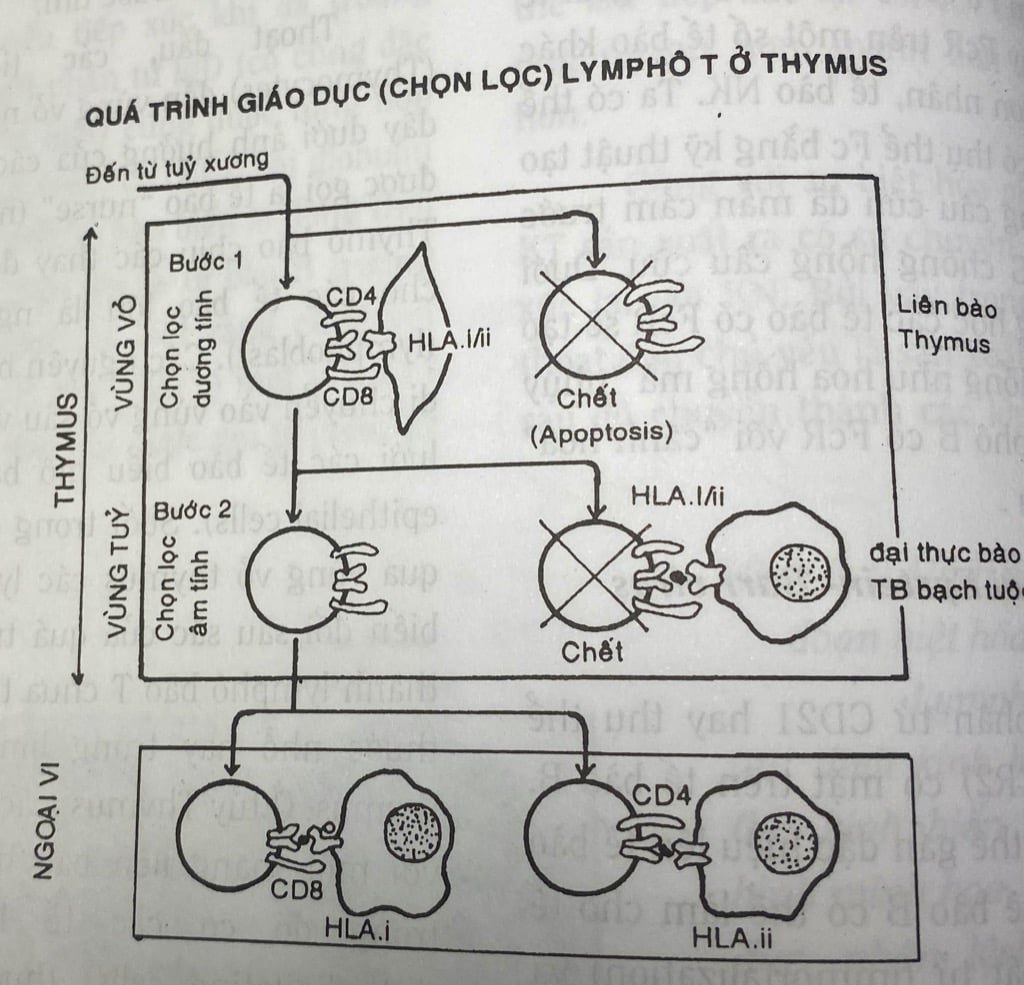
Quá trình chọn lọc trên được xảy ra qua hai bước:
Bước 1: Chọn lọc dương tính (positive selection):
Xảy ra trước và ở vỏ , tuyến Thymus. Trong bước này chỉ các tế bào T có khả năng nhận diện kháng nguyên phù hợp mô của bản thân mới được tăng sinh, nếu không có khả năng này sẽ bị chết theo chương trình (apoptosis).
Bước 2 Chọn lọc âm tính (negative selection):
xảy ra ở vùng tủy tuyến Thymus. Trong bước này các tế bào T nào có khả năng nhận diện kháng nguyên bản thân trình diện trên kháng nguyên phù hợp mô cũng của bản thân sẽ bị loại bỏ (hoặc chết, hoặc bất hoạt).
Qua 2 quá trình trên các tế bào T còn lại không còn khả năng nhận diện và phản ứng lại kháng nguyên bản thân được trình diện trên kháng nguyên phù hợp mô của bản thân và do đó tạo nên sự dung nạp ở trung ương. Sự lệch lạc trong quá trình chọn lọc này có thể là một trong nhiều nguyên nhân của một số bệnh tự miễn. Sau quá trình chọn lọc 2 bước, các tế bào T này mới thực sự trưởng thành và được di chuyển đến các cơ quan lymphô ngoại vi.
Sự hình thành các dấu ấn của tế bào T
Trong quá trình trưởng thành các tế bào T có nhiều thay đổi về thuộc tính sinh học và chức năng mà ta có thể nhận biết nhờ các dấu ấn của sự biệt hóa (differentiation markers). Trong các dấu ấn đó, thường sử dụng nhất là các dấu ấn ở bề mặt tế bào (surface markers) mà có thể dễ dàng nhận biết nhờ các KHÁNG THỂ đơn clone đặc hiệu.
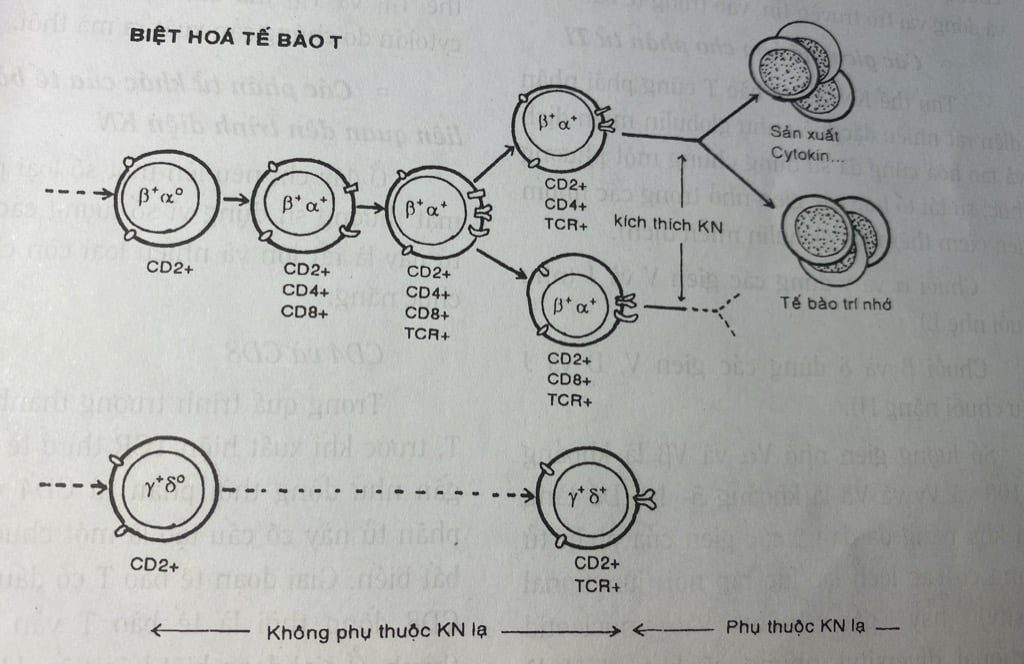
Thụ thể đặc hiệu KHÁNG NGUYÊN của tế bào T (TCR: T cell Receptor)
TCR là dấu ấn quan trọng của tế bào T và | tương đương với BCR đối với tế bào B. TCR là một phức hợp bao gồm phân tử nhận diện KHÁNG NGUYÊN gọi là Ti liên kết chặt chẽ (song không liên kết đồng hóa trị) với phân tử CD3 bao gồm nhiều chuỗi polypeptid bất biến. Như vậy có thể tóm | tắt: TCR = (Ti + CD3).
Sở dĩ gọi là Ti vì hồi đầu phân tử này được nhận biết nhờ các KHÁNG THỂ đơn chôn đặc hiệu với quyết định idiotip của phân tử trên tế bào T (T= tế bào T, i = idiotip).
Ti bao gồm hai chuỗi polypeptid, các chuỗi polypeptid này cũng có cấu tạo thành các búi (loops) nhờ các cầu nối S – S, mỗi chuỗi có hai búi và do đó cũng tạo ra 2 đô men. Bởi vậy Ti cũng thuộc trong gia đình của các phân tử có cấu trúc kiểu globulin miễn dịch. Ngày nay người ta biết được có hai kiểu cấu tạo thành Ti phụ thuộc vào bản chất chuỗi polypeptid: hoặc op (chiếm > 90% tế bào T) hoặc rò (chiếm <10%). Nên nhớ rằng mỗi tế bào T chỉ biểu lộ một kiểu duy nhất mà thôi.
CD3 bao gồm 5 chuỗi polypeptid bất biến, chúng có mặt trên mọi tế bào T trưởng thành và đóng vai trò truyền tin vào trong tế bào.
Các gien mã hóa cho phân tử Ti
Thụ thể KHÁNG NGUYÊN của tế bào T cũng phải nhận diện rất nhiều đặc hiệu như globulin miễn dịch và tạo hóa cũng đã sử dụng chung một phương thức: sự tái tổ hợp các gian nhỏ trong các nhóm gen (xem thêm bài globulin miễn dịch).
Chuỗi α và γ dùng các gien V và J (như chuỗi nhẹ L).
Chuỗi B và 6 dùng các gien V, D và J (như chuỗi nặng H).
Số lượng gian nhỏ Vα và Vβ là khoảng 50- 100 và VY và Vô là khoảng 5- 10. Để tăng thêm khả năng đa dạng, các gian của phân tử Ti cũng có các lệch lạc lúc ráp nối (junctional diversity) hay gắn thêm vào nucleotid (insertional diversity) nhưng có khác biệt là người ta không thấy các đột biến xảy ra sau kích thích bởi KHÁNG NGUYÊN lạ ở các gian này. Các quy luật về loại trừ allele cũng xảy ra với các gian của phân tử Ti như với phân tử globulin miễn dịch.
Các dưới quần thể tế bào TH và đặc điểm sản xuất cytokin của chúng
Tế bào TCD4 khi được hoạt hóa trở thàn, Tế bào hành sự (effector cells), chúng được chia thành 2 dưới quần thế là TH và TH khác – về hai nhóm cytokin được chúng sản xuất ra. Dưới quần thể Tại sản xuất chủ yếu là IL-2. INF-γ và TNF-β có chức năng chủ yếu đối với | miễn dịch qua trung gian tế bào như phản ứng quá mẫn muộn, hoạt hóa các tế bào Tc. Còn dưới quần thể TH2 có chức năng chủ yếu là giúp đỡ để hoạt hóa tế bào B tức là giúp cho đáp ứng miễn dịch dịch thể đặc hiệu. Cytokin được TH2 tiết ra là IL-4, 5, 6 và 10. Riêng IL-10 còn có tác dụng ức chế đáp ứng miễn dịch tế bào. Điều cần biết là hiện nay người ta chưa biết có dấu ấn bề mặt nào để phân biệt giữa 2 dưới quần thể Thi và TH2 mà chỉ dựa vào đặc điểm của cytokin do chúng sản xuất ra mà thôi.
Các phân tử khác của tế bào T có liên quan đến trình diện kháng nguyên
Ở đây chỉ nêu lên một số loại phân tử bề mặt thường sử dụng vì số lượng các loại phân | tử này là rất lớn và nhiều loại còn chưa biết rõ chức năng.
CD4 và CD8
Trong quá trình trưởng thành của tế bào T, trước khi xuất hiện TCR thì ở tế bào T đã có gần như đồng thời phân tử CD4 và CD8. Hai phân tử này có cấu tạo là một chuỗi polypeptid bất biến. Giai đoạn tế bào T có dấu ấn kép CD4 CD8 đồng thời là tế bào T vẫn chưa trưởng thành. Ở giai đoạn biệt hóa tiếp, khi tế bào T đã | biểu lộ TCR, tế bào T chia làm 2 dưới quần thể chỉ còn lại một dấu ấn là CD4 hoặc CD8 mà thôi (Ti+ CD3+ CD4+ CD8- hoặc Ti+ CD3+ CD4- CD8+) và lúc này là tế bào T đã trưởng thành.
– Phân tử CD4 nhận diện một vùng bất | biến của phân tử phù hợp mô lớp II và do đó tế | bào TCD4+ chỉ nhận diện KT trình diện tới phân tử HLA lớp II. Phân tử CD4 là dấu ấn của tế bào T giúp đỡ cảm ứng. Ngoài ra CD4 còn là thụ thế của HIV. | Phân tử CD8 nhận diện một vùng bất biến của phân tử HLA lớp I, do đó tế bào TCD8+ chỉ nhận diện KN trình diện qua phần tử HLA lớp I. Tế bào TCD8+ thuộc dưới quần thể tế bào T gây độc (Cytotoxic Tcell: Tc) và tế bào T trấn áp (Suppressive T cell, T suppressor: Ts).
Các phân tử liên quan đến sự bám dính (adhesion molecules)
Phân tử CD2
Trong quá trình biệt hóa của lymphô T, phân tử CD2 biểu lộ rất sớm và có vai trò như thụ thể của hồng cầu cừu nên người ta đã lợi dụng tính chất này để nhận biết lymphô T bằng phản ứng tạo hoa hồng với hồng cầu cừu (E rosette). Về chức năng sinh học, phối tử tự nhiên của phân tử CD2 chính là phân tử LFA-3 (hay CD58) có mặt trên nhiều loại tế bào. Chức năng của phân tử CD2 có thể là thụ thể truyền tín hiệu hoạt hóa hay tự hủy tùy hoàn cảnh ở một giai đoạn biệt hóa sớm của tế bào T khi chưa có sự biểu lộ TCR.
Phân tử LFA- 1. (Leucocute function associated antigen-1)
Khác với CD2, CD4, CD8, phân tử LFA – 1 có mặt trên nhiều loại tế bào. Phân tử LFA-1 là một glycoprotein gồm hai chuỗi polypeptid và có khả năng dính với một phân tử khác có tên là phân tử ICAM-1 (intercellular adhesion moleculel) cũng có mặt trên nhiều loại tế bào khác. Thông qua các phân tử bám dính này làm cho tương tác giữa tế bào T và các tế bào khác vững chắc hơn, sự dẫn truyền tín hiệu hoạt động tốt hơn.
CÁC TẾ BÀO TRÌNH DIỆN KHÁNG NGUYÊN (APC: ANTIGEN PRESENTING CELLS)
Tế bào trình diện kháng nguyên cho lymphô T
Các tế bào APC thuộc dòng tủy biệt hóa để cho các tế bào đơn nhân (hay còn gọi là các đơn nhân thực bào). Các tế bào này lưu hành trong máu rồi di trú tới một số cơ quan và tổ chức để trở thành những tế bào có | tên gọi khác nhau và chức năng cũng khác nhau một phần.
Ví dụ:
- Tổ chức bào: ở các tổ chức.
- Đại thực bào: ở phúc mạc, màng phổi, phối, lách.
- Tế bào Langerhans: ở thượng bì. BỘT
- Tế bào Kupffer: ở gan. Tế bào bạch tuộc: ở hạch bạch huyết.
- Tế bào xòe ngón tay: ở hạch bạch huyết, Thymus.
- Tế bào thần kinh nhỏ (microglial cells) ở não…
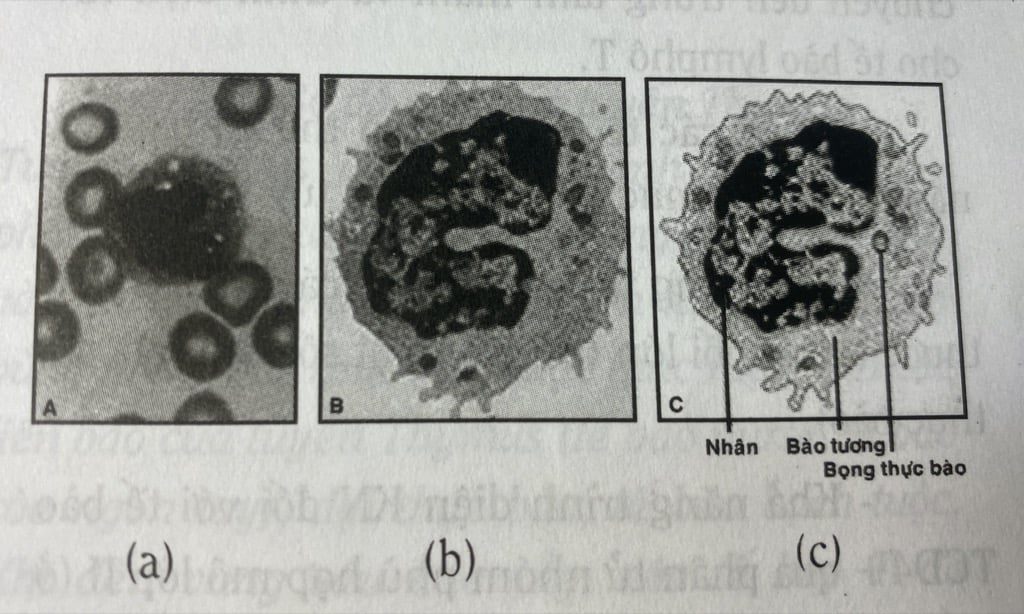
Bởi vậy các tế bào trình diện kháng nguyên là một quần thể tế bào dị đồng. Có thể nêu ra một số tế bào thuộc nhóm này cho thấy chúng có phương thức hoạt động không giống nhau:
Các tế bào Langerhan ở trong da, sau khi bắt giữ KN sẽ di chuyển theo đường bạch mạch đến vùng cận vỏ của hạch bạch huyết. Các tế bào này có hình dạng khác trước: có các tua dài đế tương tác với các lymphô T (bản chất là hoạt động trình diện KN) nên chúng có tên gọi là tế bào xòe ngón tay. Các tế bào này giàu các phân tử HLA lớp II, trình diện KN hiệu quả.
Các tế bào bạch tuộc có mặt trong nang lymphô B nguyên thủy và thứ cấp ở hạch, ở lách, ở tổ chức lymphô dưới niêm mạc. Các tế bào này không di chuyển mà tạo thành một mạng lưới tại chỗ. Chúng hầu như không biểu hiện phân tử HLA lớp II do đó không trình diện với tế bào lymphô T mà “trình diện” phức hợp KN cho tế bào lymphô B thông qua thụ thế bố thể và thụ thể Fc. Thực ra đây không phải là kiếu trình diện KN theo ý nghĩa kinh điển. Gần đây người ta phát hiện trong nhóm này có một loại tế bào được đặt tên là tế bào bạch tuộc của trung tâm mầm (germinal center dendritic cells – GCDC) có biểu lộ phân tử HLA lớp II, chúng di chuyển đến trung tâm mầm và trình diện KN cho tế bào lymphô T.
Tất cả các tế bào trên có thể có hại khả năng nhưng ở mức độ khác nhau tùy loại, đó là:
- Khả năng thực bào kể cả các hạt có kích thước tương đối lớn (bởi vậy một số có tên là đại thực bào).
- Khả năng trình diện KN đối với tế bào TCD4+ qua phân tử nhóm phù hợp mô lớp II (ở người là nhóm HLA lớp II).
Nhờ sự phân bố ở khắp các vị trí trọng yếu thường bị xâm nhập bởi các vi sinh vật như diện tiếp xúc (da, niêm mạc, các khoang..) ở tuần hoàn (gan, lách…) nên chúng có vai kiểm soát trong phòng vệ tuyến đầu. Hơn nữ các đơn nhân thực bào còn có khả năng : diệt những tế bào biến đổi của cơ thể (kể cả bào ung thư) bằng thực bào hay sản xuất ra các cytokin (IL1, TNF, IFN…). Khả năng bắt giữ các vi sinh vật, các tế bào bị biến đổi có được là nhờ các thụ thể ở bề mặt của đơn nhân thực bào (kể cả thụ thể với phần Fc của globulin miễn dịch và C3b của bổ thể).
Như vậy các tế bào thuộc nhóm đơn nhân thực bào tham gia vào trong đáp ứng miễn dịch cả phần cảm ứng (afferent or induction limb) lẫn phần hành sự (efferent or effector limb).
Về phân loại, các tế bào trình diện KN thuộc loại các tế bào miễn dịch không đặc hiệu song lại có vai trò quan trọng trong trình diện KN để mở đầu cho một đáp ứng miễn dịch đặc hiệu cho nên người ta còn gọi các tế bào này là các tế bào giao diện giữa miễn dịch không đặc hiệu và miễn dịch đặc hiệu.
Tế bào trình diện KN cho lymphô B: Tế bào bạch tuộc ở nang (FDC: Follicular Dendritic Cell)
Các tế bào FDC hiện diện ở các nang nguyên thủy của hạch lymphô, dường như chúng có vai trò trong việc giữ lại các lymphô B tại đây và tham gia chính trong quá trình chọn lọc và thuần thục của đáp ứng miễn dịch dịch thể.
Nguồn gốc của tế bào FDC còn chưa rõ nhưng người ta biết rằng nó không giống như các tế bào bạch tuộc trình diện KN cho tế bào T ở chỗ: chúng không có nguồn gốc từ tế bào tủy xương, không biểu lộ các phân tử nhóm phù hợp mô lớp II, không thực bào và xử lí KN mà nó chỉ bắt các KN nhờ các thụ thể (với các thành phần bố thế) CR1, CR2, CR3 và thu thể E. Các KN được bắt giữ qua các thụ thế này vẫn ở nguyên trạng trên bề mặt tế bào một thời gian dài vài tháng có khi đến vài năm. Các KN ở trên bề mặt tế bào FDC có chức năng trình diện cho các lymphô B có các thụ thể đặc hiệu. Các lymphô B đặc hiệu sẽ được tăng sinh, hoạt tác và với sự giúp đỡ của lymphô T để biệt hóa thành các tương bào sản xuất các KT đặc hiệu còn một bộ phận trở thành tế bào trí nhớ. Về hình thái vị thế, khi có sự tăng sinh, hoạt hóa và biệt hóa của lymphô B các nang nguyên thủy trở thành các trung tâm mầm.